Division
Forestry Services




Burn Ban Map: Click on individual counties in yellow to view/download current burn ban resolutions. Burn Ban FAQ.
OK Situation Report: Sign-up to receive the Oklahoma Fire Situation Report by email or text.
The Role of Fires in Oklahoma Landscapes
Many of Oklahoma’s landscapes have evolved with fire as a natural and necessary contributor to their overall health and renewal. Many plant species require fire to germinate, to establish, or to reproduce, or all three. Native Americans used fire across Oklahoma to provide better access, improve hunting, and ridding the land of undesirable species so they could farm. Early European settlers to our state observed this and continued the practice of using fire as a beneficial agent.
But as more settlers arrived, an encroaching urban interface and losses to timber, farm and range land called attention a growing wildfire problem and led to fire being labeled as destructive. Since that time great effort has been made to exclude fire from the landscape. But removing fire from the landscape has had consequences. Many of our lands are no longer healthy and the growth which has continued in the absence of fire resulted in an accumulation of fuels increasing the overall risk of wildfires.
As knowledge accumulated, the use of “prescribed” fire grew and natural resource professionals now include fire as an appropriate tool to manage forests, woodlands and range. Properly used, fire remains an excellent tool for restoring and managing many Oklahoma landscapes.
Learn more about fire’s role in Oklahoma ecosystems and the proper use of prescribed fire. Watch our video A Land in Balance.
Wildland fires are a force of nature that can be nearly as impossible to prevent, and as difficult to control, as hurricanes, tornadoes, and floods. Wildfire can be a friend and a foe. In the right place at the right time, wildland fire can create many environmental benefits, such as reducing grass, brush, and trees that can fuel large and severe wildfires and improving wildlife habitat. In the wrong place at the wrong time, wildfires can wreak havoc, threatening lives, homes, communities, and natural and cultural resources.
Report a Wildfire
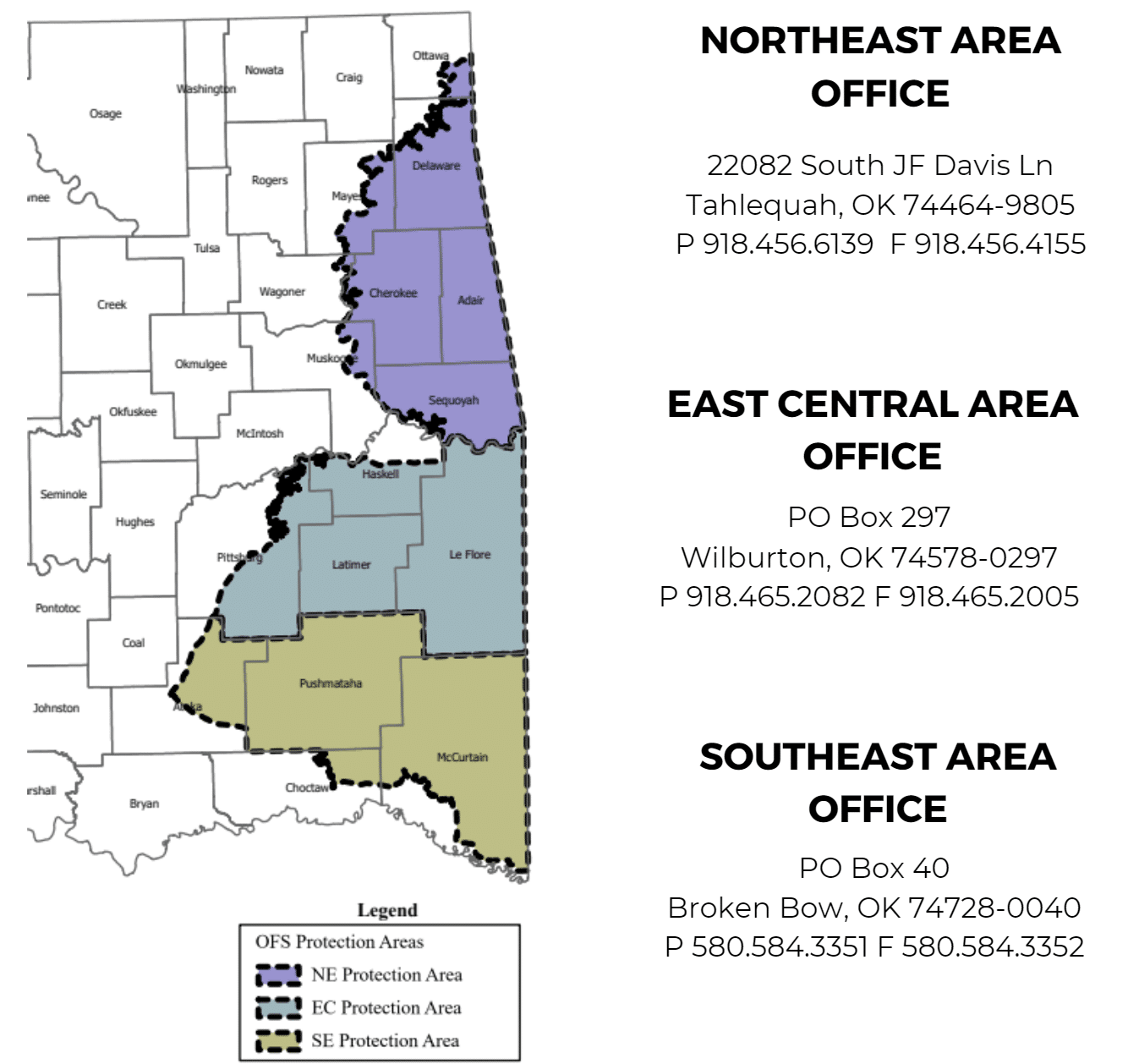
In the OFS designated initial attack area (see map) call:
- Northeastern Oklahoma: 1-800-299-3473
- East Central Oklahoma: 1-800-375-2056
- Southeastern Oklahoma: 1-800-299-2468
For all other areas in Oklahoma, report fires to the nearest fire department or CALL 911.
Report Arson
1-866-NO ARSON (866-662-7766) or to your nearest OFS forester.
Oklahoma Forestry Services is both honored and pleased to serve alongside of and assist the many fire departments in our state. In addition to providing direct assistance during ongoing incidents we work to improve fire department capacity through our Community Fire Assistance Program by providing funding opportunities, equipment and training in addition.
From the pine forests of the east, to the unique Cross Timbers forest of central Oklahoma and the riparian areas of the Great Plains, OFS Foresters work with landowners who want to actively manage and care for their trees and property. No matter what your goals might be, your OFS Forester is there to provide you the best management keys to success.
WATER QUALITY
TREE AND FOREST HEALTH
FOREST MARKETS AND PRODUCTS
URBAN AND COMMUNITY FORESTRY
FOREST LEGACY
OKLAHOMA FOREST ACTION PLAN
FOREST MANAGEMENT GRANT/COST-SHARE PROGRAMS
PLANNING YOUR FOREST’S FUTURE
FOREST TAXATION AND ESTATE PLANNING
Click Here for a printable list of downloadable list of Consulting Foresters of Oklahoma.
Oklahoma Sawmill Locations:
If you would like your sawmill added to the map, please complete the form found here.
OFS provides technical assistance to plant, maintain, and restore the health of your trees and the trees of your community. Communities have unique and valuable forests, growing along streets, greenbelts, parks and our own backyards. Urban forests enhance our quality of life in numerous ways:
- Cleaning the air we breathe
- Raising real estate values
- Ensuring high quality drinking water
- Modifying temperature extremes
- Reducing noise pollution
- Improving the aesthetics of Oklahoma’s landscape
2024 Urban and Community Forestry Assistance Grant Program
2024 State Forester Signed Letter
2024 U&CF Assistance Grant Program Application (download for fillable PDF)
State Nursery and Tree Improvement Center
Oklahoma Forestry Services has a long history of forest tree improvement and seedling production. This commitment continues today as we strive to provide the highest quality seedlings available anywhere.
OFS operates two facilities which specialize in forest regeneration: the Forest Tree Improvement Center (FTIC), located in Idabel, and the Albert Engstrom Forest Regeneration Center (State Nursery), located in Goldsby. The goals of Oklahoma’s Forest Regeneration Program are to keep forestlands in forest, and to offer tree planting as a viable solution to some of Oklahoma’s most pressing environmental issues.
Ordering Seedlings
OFS sells over 30 varieties of trees and shrubs, chosen for their ability to meet the challenges of Oklahoma’s tough climate. Years of selecting from the best seed sources available for each species and careful growing practices in our nursery have resulted in seedlings adaptable to a wide range of planting conditions. Talk to our Regeneration Staff or your local Service Forester for recommendations for your particular site and how to properly plant seedlings.
Seedlings are distributed as bare-root and containerized and can be planted for a number of different purposes including timber production, wildlife habitat improvement, erosion control and windbreaks. The State Nursery also has special options available including Genetically Improved Southern Pines and Conservation Planting Packets.
Place an order for seedlings starting in October and they are shipped in January through March of each year. If you have questions regarding seedling orders or policies, please contact the State Nursery at 405-288-2385 or 800-517-3673. Or send us an email.
See the latest catalog and order seedlings here:
Not all tree species are ideal for all areas of the state. OFS is offering a list of native tree species by county for use by Oklahoma landowners as a tool to help choose the best trees to plant in their respective areas. It can be downloaded here as an Excel file, a Google Tables file, or as a PDF. If you’d like to purchase a copy of our Forest Trees of Oklahoma book, you can do that here.
How to plant seedlings:
OFS foresters are available at no cost to help assist you with any tree planting questions. We also offer a brochure on how to plant a seedling by hand. This video also has helpful information on how to plant your seedlings.
Work at the Nursery
OFS needs Nursery Technicians in Goldsby. For details click here.